
Int J Chem Res, Vol 6, Issue 3, 1-9Review Article
POTENTIAL OF SCORPION VENOM FOR THE TREATMENT OF VARIOUS DISEASES
MOHSIN JAVED1, SHABBIR HUSSAIN2*, MUHAMMAD AMJAD KHAN3, AFFIFA TAJAMMAL4, HIRA FATIMA1, MUHAMMAD AMJAD4, AMINA ZAHID4, MAFIA UMER4, SIDRA AMEER ALI4, MADEEHA YAQOOB4
1Department of Chemistry, University of Management and Technology, Lahore, Pakistan, 2Department of Chemistry, Khwaja Fareed University of Engineering and Technology, Rahim Yar Khan, Pakistan, 3Department of Biology, Lahore Garrison University, DHA Phase VI, Lahore, Pakistan, 4Department of Chemistry, Lahore Garrison University, DHA Phase VI, Lahore, Pakistan
*Email: [email protected]
Received: 02 Mar 2022 Revised and Accepted: 16 May 2022
ABSTRACT
The venom of various scorpion species finds significant therapeutic applications. It is rich in neurotoxins, enzymes, enzyme inhibitors, histamine, lipids and different salts from which peptides demonstrate a great potential against a variety of diseases. Many biological functions e. g., bradykinin potentiating, hemolytic, anti-cancer, anti-microbial, and anti-inflammatory potential are being regulated by non-disulfide-bonded peptides. Therefore, it is motivating to use these properties for the treatment of cancer, cardiovascular diseases, diabetes, AIDS, apoplexy, influenza H5N1, paralysis, epilepsy, malaria, measles, severe combined immunodeficiency, fever blisters and diabetes. Scorpion venom has shown the presence of 100,000 bioactive compounds but only 1 % of these have been purified, isolated and characterized by HPLC and mass spectroscopy etc. For the production of high-quality antivenom with specific antibodies, gentler electrical stimulation is a better method as compared to manual production. Recombinant DNA technology has facilitated the identification of new components. Some important medicinal compounds isolated from scorpion venom include HsTX1 (from Heterometrus spinnifer), mucroporin-M1 (from Lychas mucronatus), chlorotoxin and charybdotoxin (from Leiurus quinquestriatus hebraeus). B. leptochelys venom has shown the presence of at least 148 components. Six novel long-chain peptides were isolated from the scorpion Buthus martensi Karsch venom. Crude venom of L. Abdullah bayrami displays a proliferative effect on MCF-7 cells and also shows antimicrobial potential. A new toxin derived from the venom of Liocheles waigiensis [U1-liotoxin-Lw1a (U1-LITX-Lw1a)] displays significant insecticidal action. The computational studies may play an important role while developing ion channel drugs from venom peptides.
Keywords: Scorpion venom, Toxins, Cell channels, Disease treatment, Drug formation
© 2022 The Authors. Published by Innovare Academic Sciences Pvt Ltd. This is an open access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/)
DOI: http://dx.doi.org/10.22159/ijcr.2022v6i3.204. Journal homepage: https://ijcr.info/index.php/journal
INTRODUCTION
Scorpions are famous all over the world for their poisonous sting, which is highly painful and causes a lot of health issues, including severe pain, inflammation, hypertension, cardiac arrhythmia; it may also lead towards muscle paralysis or even death in children. Their venom has been reported to show significant hazardous effects on health [1] as it inhibits the functioning of Na+/K+ ATPase pump and may cause the failure of the nervous system [2]. Management of poisoning is one of the biggest challenges faced by health care professionals globally [3]. There are reports of the treatment of numerous diseases through the venom of some poisoned species e. g., lionfish spine poison extract [4] and snake venoms [5-7]. Scorpions venom also finds applications in the diagnosis of various diseases, including tetanus, epilepsy, chronic and acute convulsion [8]. Scorpions (about 1500 species) are known as one of the ancient species which have been living on the earth since 400 million years ago. They are the causes of approximately 1.2 million stings and 3250 human deaths every year throughout the world [9]. In Saudi Arabia (especially in the northwest region), there is highly prevalence of scorpion stings in Riyadh [10]; about 25 species of snakes are proved to be highly toxic in this country [11]. The most toxic venom belongs to the genus “Androctomus,” which also exists in KSA [12]. The common outdoor activities and hot climatic conditions result in the common exposure of local residents to scorpion attacks [10].
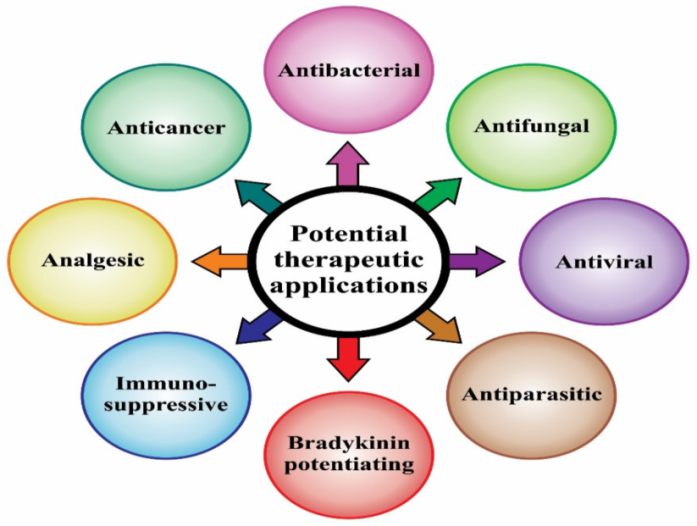
Fig. 1: Potential therapeutic application of scorpion venom [13]
The scorpion venom can interfere with the ion channel operating mechanisms in the body and result in the blockage of the normal neurotransmitter signaling path. Despite of its negative effects, it is also beneficial for human beings as it can be used for the cure of various deadly diseases (fig. 1) [13], including cancer, [14] blood and pancreatic cancers [15] and is also useful for the synthesis of toxin-neutralizing vaccine [16]. It has also been reported that the scorpion venom selectively attacks on cancerous cells without showing any interaction with healthy cells. Its application results in the rupture of normal architecture of cancerous cells and also decreases cell proliferation [17].
Current studies were performed to review the chemical nature of scorpion venom and its potential therapeutic applications.
MATERIALS AND METHODS
Review studies were performed on the chemical nature [18-26] and therapeutic potential [8, 17, 27-72] of scorpion venom [2, 9, 14] and relevant literature is reviewed from 1993 to 2022.
RESULTS AND DISCUSSION
The venoms of many animals (scorpion, frog, toad, snake, etc) and their active components (protein/non-protein toxins, enzymes, peptides, etc) possess anticancer potential [18]. Various bioactive components (such as peptides, nucleotides, amino acids and lipids) of scorpion venom may cause severe systemic inflammation after they are injected into human beings [19]. Toxin peptides of scorpion venom greatly interact with the normal functioning of excitable/non-excitable cells; they can modify or block the ion-channel functions and thus find applications to control cell excitability. Many venom proteins find potential applications as antibiotics. The deleterious effects of venom on human beings can be neutralized by venom itself. Moreover, venom has also been found effective in the production of many antibodies in experimental animals (sheep and horse) [19]. The toxicity of scorpion venom is governed by some factors i.e. scorpion’s age, genus and species, physiology, region, and feeding habits [20]. There is a very critical procedure to collect high-quality venom from scorpion. By abdomen stimulation, less toxic and transparent venom is produced but when the stimulation is done externally through electric shock the scorpion generates very concentrated toxin [18]. Both kinds of venoms (less toxic and concentrated) differ from each other in physical and chemical properties as the latter contains a high concentration of salts which greatly affect the potassium ions of the targeted creature [20].
Scorpion finds a fantastic potential against various diseases and is used to synthesize vaccines which have ability of neutralizing several toxins. Novel methods and separation techniques have been adopted to characterize venom components; these components are highly valuable for the development of medicinal drugs. Scorpion venom is constituted of small neuro-toxic peptides, amino acids, enzymes, enzyme inhibitors, histamine, lipids and different salts [21]. Approximately 800 natural protein toxic components were listed in data banks of scorpion venoms [22].
Chemical nature of scorpion venom
The symptoms associated with envenomation are owed to the presence of a large number of bioactive components such as amino acids, nucleotides, lipids and peptides (most important component) in scorpion venom [19]. The toxin peptides of scorpion venom can be categorized into two main classes (i) disulfide-bonded peptides and (ii) non-disulfide-bonded peptides [23]. Disulfide-bonded peptides consist of 3 to 4 disulfide bridges and are generally classified according to their mode of interaction with different ion channels (Ca2+, Na+, Cl, and K+) of cells. In mammalian organisms, these membrane-bound ion channels play an important role in regulation of the normal cell behavior. Many biological functions such as hemolytic, bradykinin potentiating, anti-inflammatory, anticancer, and anti-microbial activities are regulated by non-disulfide-bonded peptides [24]. The scorpion toxins have the ability to identify and bind to the ion channels. Toxins vary in the length of their amino acid chains depending upon the nature of metal channels to which they have the ability to recognize. The toxins generally contain 29 to 41 amino acids which are specific for the interaction with K+and Cl-channels. Toxins having the recognition ability of Na+channels are 60 to 76 amino acids longer (4 disulfide bridges), while the toxins interacting with Ca+channels are rare and have variable lengths of their amino acid residues. However, these toxins are very much similar to each other in terms of their main 3-dimensional folding pattern and primary sequence. The central core of all toxins is generally formed by the analpha-helix segment and many anti-parallel beta-sheet stretches [25]. β-toxins have the ability bind to sodium channels permitting an abrupt shift towards a more negative potential of the membrane. CSS IV and Ts1 (another name Tsγ) obtained from Centruroides suffuses (scorpion venom) and Tityus serrulatus (Brazilian scorpion), respectively, are β-toxins which can be bound to the sodium channels [26]. The presence of 36 amino acids and 4 disulfide bonds has been verified in the chlorotoxin peptide of Leiurus quinquestriatus venom. NMR Spectroscopy has shown the presence of 3 anti-parallel ß-sheets, which are cross-linked through 3-disulfide bonds with an ɑ-helix and the fourth one is connected to the beta-strand with a small N-terminal. The binding of metalloprotease-2 with chloride channels is blocked by chlorotoxin. Endocytosis of MMP-2 is caused by chlorotoxin; it results in the depletion of Cl-channels in the cell surface. This chlorotoxin only binds to the cancerous cells and not to the normal ones [21].
Purification and isolation of the desired component of scorpion venom
When scorpion venom is injected into the human body, it may cause a lot medical issues and even death. New biotherapeutics have been developed from scorpion venom due to the presence of a large number of bioactive molecules. The presence of several proteins, peptides, amines and nucleotides enables the scorpion venom to act as an interesting therapeutic agent against many current and emerging diseases [13, 44].
Isolation of the individual components of scorpion venom is necessary for the determination of their biological importance [73]. Scorpion venom contains a diverse range of bioactive compounds (about 100,000 in number) but only 1 % of these have been purified, isolated and characterized [74]. The separation and purification of the venom polypeptides can be performed by applying a 4-steped method; it consists of pre-treating the scorpion venom solution, reasonably selecting the protein concentration before separation and purification of scorpion venom solution, use of a buffer liquid and an eluting flow rate [75]. The introduction of recombinant DNA technology (such as transcriptome analysis), has aided in the identification of novel components; however, some components cannot be directly isolated from the venom [76]. Neurotoxins make the major proportion of scorpion venom and are responsible for the various pathological manifestations of envenoming [13]. Scorpion venom generally contains four kinds of neurotoxins depending upon the peptides which modulate calcium-, chloride-, potassium-or sodium-gated channels [77].
Peptides are generally recognized as potential therapeutics due to their high selectivity and relatively safe mode of action. Peptides purified as single compounds can be used as useful drugs at appropriate concentrations [78]. These tiny peptides are the most studied scorpion venom components especially due to the broad ranges of their pharmacological applications and diversity. Depending upon their structural features, peptides have been grouped into 3 main superfamilies:
Calcins
Peptides containing cysteine-stabilized (CS) α/β motifs
Non-disulfide bridged peptides (NDBPs) [13]
Scorpion venom also contains amines, nucleotides, free amino acids, mixtures of inorganic salts, lipids and enzymes [21]. However, a number of technological hurdles are needed to be overcome before the commercialization of venom-derived biotherapeutics [13].
A comparison of two approaches (manual and electrical stimulation of adult scorpions) for collecting scorpion venoms on a wide scale has been published. It was found that high-quality antivenom with specific antibodies can be produced by using the gentler electrical stimulation method. Electrical stimulation is used to acquire scorpion venom more quickly and in higher quantities; this procedure also produces venom free from hemolymph contaminants e. g., hemocyanin. However, scorpions suffer from maximal trauma and consequently hemocyanin secretion during manual obtainment [79]. With the improvement of technology over the past century, the separation of venoms and full characterization of the individual peptides and proteins has become very easier. A typical approach for isolating and evaluating peptides and other bioactive substances from venomous animals is shown in fig. 2 [78].
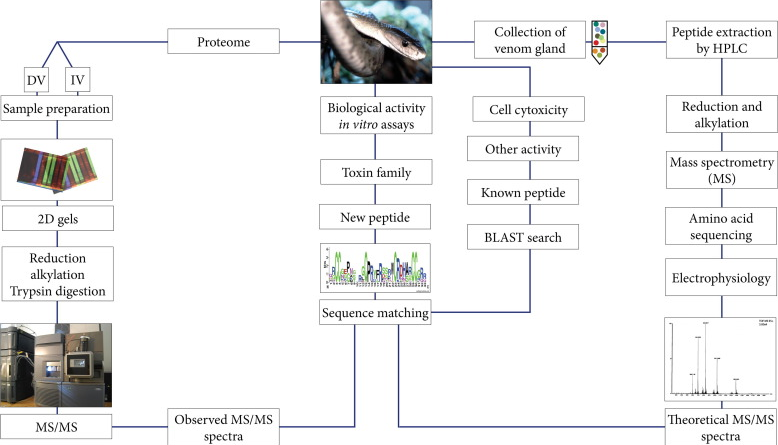
Fig. 2: A typical workflow for isolation and screening of peptides and other bioactive compounds from venomous animals e. g., the black mamba (Dendroaspis polylepis) [78]
The venom of the deathstalker scorpion (Leiurus quinquestriatus hebraeus) can be screened for chlorotoxin and charybdotoxin. Chlorotoxin is one of the venom's most prominent peptides, with 36 residues and four disulfide bonds; it possesses the property of blocking chloride channels and is effective against cancer (fig. 3) [78]. Charybdotoxin is a Ca2+-activated K+channel inhibitor that is also active against KV1.3 [78, 80]. The margatoxin produced by Central American bark scorpions (Centruroidesmargaritatus) is more than 20 times more effective than charybdotoxin against KV1.3 (Ki 50 pM) and has no effect on Ca2+-activated K+ channels [78, 81]. The margatoxin was found to be effective against delayed-type hypersensitivity (DTH) in a mini-pig model [78, 82].
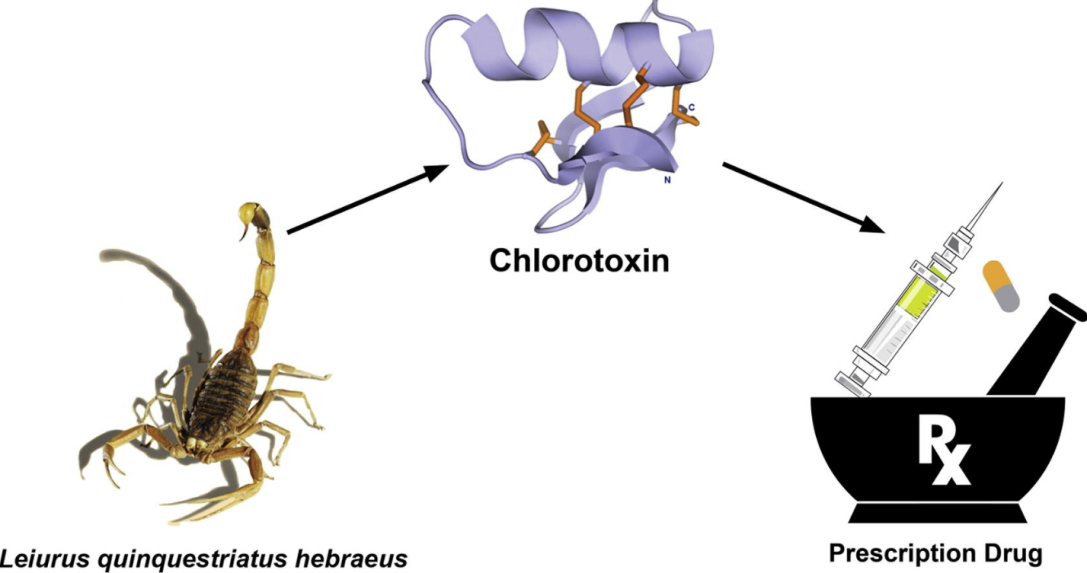
Fig. 3: Medicinal use of chlorotoxin [78]
HsTX1, a 34-residue peptide containing an uncommon fourth disulfide link, was discovered in the venom of the scorpion Heterometrus spinnifer. It has the ability to block KV1.3 channels [78, 83] and is relatively selective versus KV1.1 [78, 84]. It is worth noting that KV1.3 blockers have emerged as promising tools for autoimmune diseases [78, 85]. 148 components have been analyzed by LC/MS performed on venom of the B. leptochelys. Through bioassay-guided HPLC fractionation, the four peptides (Bl-1, Bl-2, Bl-3, and Bl-4) were isolated [86]. The peptide profile of the Leiurus abdullahbayrami venom (one of the deadliest venoms among Turkish scorpions) was determined by mass spectroscopy, reversed-phase chromatography, size-exclusion and electrophoretic methods. Their cytotoxic and antimicrobial activities were also assessed against breast cancer cell line (MCF-7) and several bacterial/fungal species, respectively. About half of the dry weighed venom of L. Abdullah bayrami crude is made up of proteins. The presence of 6 to 7 kDa peptides was detected using microfluidic capillary electrophoresis. Mass spectroscopy has shown the identification of 45 unique peptides which have masses between 1 to 7 kDa (29% of 1-2 kDa and 31% of 3-4 kDa). Due to high concentrations of polyamines, calcium and potassium ions in spider venoms, the crude venom shows a proliferative effect on MCF-7 cells. The venom also exhibited antimicrobial potential against the targeted gram-negative bacteria [87]. Two-dimensional mixed-mode reversed-phase chromatography coupled with tandem mass spectrometry has shown the presence of 6 novel long-chain peptides in the venom of scorpion Buthus martensi karsch [73]. A new toxin [U1-liotoxin-Lw1a (U1-LITX-Lw1a)] was isolated from the scorpion Liocheles waigiensis. It is the first example of scorpion-venom peptides adopting a fourth structural fold and has shown significant insecticidal activity [88].
The action of toxins on various ion channels
It has been reported that many toxins can change the normal functioning of K+ channels. It is proposed that computational studies may play a crucial role in developing ion channel drugs from venom peptides [29, 30]. Many short-chain peptides are blockers of potassium ion channels and cause immune suppression [29, 31], while long-chain peptides are blockers of sodium ion channels and can treat pain [32].
Martentoxin, a pure 37-amino acid toxin was extracted from Chinese scorpion venom. It can block large-conductance Ca2+-activated K+(BK) channels in chromaffin cells in the adrenal medulla. Martentoxin at a dose of 100 nM has the ability to strongly block the BK channels of chromaffin cells in the adrenal medulla. The BK currents blocked by martentoxin can be recovered much faster as compared to those caused by charybdotoxin [33, 89]. By reversible blockage of the K+ channels, the toxins of Brazilian scorpions (such as T. stigmurus, T. bahiensis, and T. serrulatus) can limit T cell proliferation and IL-2 production [34-36]. In scorpion venom, a lot of enzyme inhibitors were also reported, which help to understand the biological functions of scorpion toxins. Specific toxins present in Chinese scorpion species play an important role as protease inhibitors, antimicrobial peptides, K+ channel blockers, and Na+ channel modulators [37]. The propagation and generation of action potentials are dependent on sodium channels [38].
There were investigations on the peptides that can recognize Ca+2-, K+-or Na+-channels of excitable cells. The most important are the peptides specific for Na+-channels due to their medical relevance; they are more than 300 in number. They can detect mammalian Na+-channels and alter the opening/closing kinetic mechanisms of ion channels and thus cause anomalous depolarization of excitable cells. Peptides that target K+-channels act as pore blockers. Although they are not life-threatening, yet they block potassium permeability, cause many electrophysiological issues (as well as discomfort) and are involved in many intoxication symptoms. More than 140 K+ -channels have been reported till now. The scorpion venom also consists of calcins (peptide toxins), including opicalcin 1 and 2, hadrucalcin, hemicalcin, maurocalcin and imperatorin, which can recognize Ca+2-channels. Some of these calcins have gained popularity in designing novel drugs because they show direct interaction with ryanodine receptors. The harmful effects of these 3 peptides can be neutralized by using specific anti-venoms e. g., immunoglobulins prepared from horses [27]. Fig. 4 displays the mechanism of K+channel activity in open/closed state as well as N-or C-type inactivation [71].
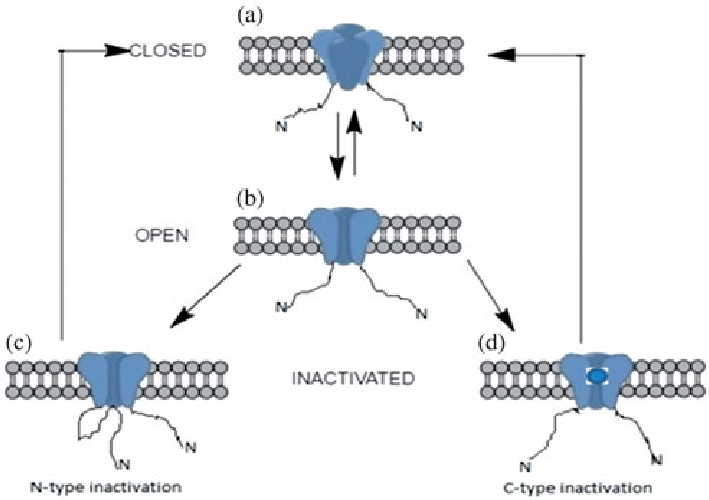
Fig. 4: Mechanism of K+channel activity (a) closed state (b) open state (c) N-type inactivation (d) C-type inactivation [71]
Potential against the proliferation of cancerous cells
There have been claims that scorpion venom can be used to treat cancer [41]. Scorpion venom is a complex mixture of peptides and proteins, most of which are neurotoxins. These toxins can bind and modulate multiple ion channels (Ca2+, Cl−, K+ and Na+) in excitable and non-excitable tissues [28]. The characteristic feature of these peptides is to decrease cell proliferation and apoptosis and also to inhibit many signaling processes which result in cancer [14, 21]. Kv expression and apoptosis are strongly linked to potassium ion channels [42]. The medical significance of venom is owed to the presence of a broad spectrum of ion channel toxins. Some animal venoms were successfully applied to treat breast cancer [43]. There are reports for the treatment of thousands of cancer cases through the blue scorpion venom (endemic to Cuba). The cancer cells are attacked by the protein chain which is present in the blue scorpion venom [45]. Fig. 5 displays the mechanism of inhibition of the growth of cancer cells by scorpion venom [71].
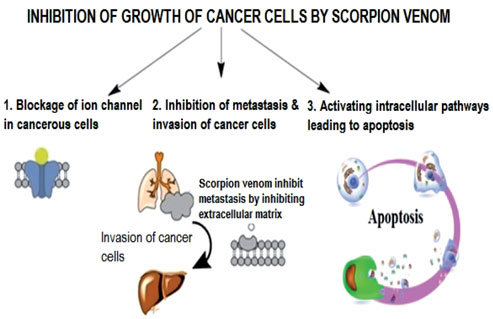
Fig. 5: Mechanism for inhibition of growth of cancer cell by scorpion venom [71]
Various scorpion species in cancer treatment
Blue (or red) scorpion (Rhopalurus junceus) is famous due to its antineoplastic activity in the Dominican islands and Cuba. Its venom maintains energy in cancer patients and also acts as a pain reliever [44]. The protein present in it can inhibit the proliferation and growth of cancer cells [43, 44]. The venom of the Blue or Red Scorpion (Rhopalurus junceus) also reduces the intensity of pain and restores energy in cancer patients. The venom extract of Blue Scorpion can behave as an anti-inflammatory, analgesic and anti-cancer agent [45]. The venom of Tityus discrepans scorpion contains two peptides, namely neopladine and neopladine, which cause apoptosis in human breast cancer cells and show marked defects [46]. The venom of Odontobuthus doriae has proteolytic enzymes. It possesses lactase dehydrogenase (LDH), which is a cytotoxic and apoptotic agent and can lower cell viability as it activates the caspase-3 and depolarization of mitochondria. Proteolytic and gelatinolytic proteases, which act against adenocarcinoma cell lines of human lungs are extracted from the scorpion Mesobuthus gibbosus [47]. By arresting S-phase and increasing reactive N intermediates, Odontobuthus doriae venom promotes apoptosis in human breast cancer cells [48]. Peptides present in the venom of Centruroides margaritatus cause a reduction in tumor size [49].
The cell cycle, cell proliferation, and cell growth can all be influenced by various components of scorpion venom. In Cuba, the use of Rhopalurus junceus as traditional medicine has been reported for the treatment of cancer [17]. The venom from Indian black scorpion (Heterometrus bengalensis) can induce the inhibition of K562 and U937 cell growth; it also possesses the specific characteristics of apoptosis such as DNA degradation, chromatin condensation, and membrane blebbing [8]. The venom of Leiurus quinquestriatus (Deathstalker scorpion) contains thirty-six amino acid peptides which block the chloride channels [50, 51]. Heterometrus bengalensis contains antiproliferative and apoptogenic properties against chronic myelogenous and bengaline [8]. The Buthus martensia (Chinese red scorpion) venom contains hyaluronidase (BmHYA1) which is responsible for metastasis and decreases the proliferation of breast cancer and possesses antineoplastic therapeutics with no toxic side effects [52, 53].
Potential against HIV/AIDS
One CD4 receptor and two other co-receptors (CXCR4 and CCR5) of HIV-1 can affect T cells [54]. CCR5 is considered as the major co-receptor for the transmission of HIV-1 [55]. Kn2-7 from Mesobuthus martensii scorpion was recognized as an effective anti-HIV-1 peptide; it has the ability to inhibit HIV-1 subtype CCR5-tropic, pseudotyped virus (PV) and CXCR4 tropic (NL4-3) PV strains. It was reported that the peptide Kn2-7 can protect against HIV-1 by interacting with viral components [56].
Potential against herpes simplex virus
The herpes simplex virus type 1 can infect human epithelial tissues, causing a variety of problems like blinding keratitis, encephalitis, oral mucosal lesions and meningitis [57]. HSV-1 greatly infects the sensory ganglia [58]. The venom of heterometrus petersii (scorpion) contains cationic peptides, which are effective against Herpes simplex virus type 1 infection. Both Hp1239 and Hp1036 peptides exhibit extracellular viricidal effects, morphological changes and strong inhibitory potential against HSV-1 when they are added to the infectious site. Some activities, such as viricidal activities and membrane penetration (which cause intracellular anti-viral effects) are related to amphipathic a-helix. The peptides of scorpion venom can make viral particles inactive and thus inhibit viral proliferation at the post-infection stage [59].
Potential against measles, influenza H5N1, Severe acute respiratory syndrome
RNA viruses cause diseases such as measles, influenza H5N1, and SARS-CoV, which are responsible for mortality and morbidity in children. Available treatments for measles viral infection possess varying side effects, such as anemia and teratogenicity [60]. Many approaches such as adenosine, peptide inhibitors, guanosine nucleosides, coumarins, modulators of cholesterol synthesis, brassinosteroids, and anti-sense molecules have also been failed [61].
A famous cationic peptide, namely mucroporin is found in the scorpion venom of Lychas mucronatus; it shows bacteria inhibition. Gram-positive bacteria are efficiently inhibited after the substitution of their amino acid with Mucroporin-M1 [62]. Hp1090 (a helical peptide in scorpion venom) acts as infection initiation and inhibits replication of HCV [63]. Mucroporin-M1 shows activities against bacteria and viruses. The dual antimicrobial activity of this peptide enables it to be used as a good antiseptic agent for hand/mouthwashes. Mucroporin-M1 blocks the functions of SARS-CoV, influenza, H5N1, and MeV by direct viricidal action [64].
Potential against cardiovascular diseases
Cardiovascular illnesses are the leading cause of death in modern society. Several toxins, including integrins are present in scorpion venom, which have the ability to disrupt blood coagulations. A peptide toxin present in the venom of Androctonus australis garzonii can induce the atrial natriuretic peptide secretion while the venom of Buthus martensii scorpion contains BmK I toxin, which moderates the contraction of the heart [65, 66]. The venom of Centruroides margaritatus contains a peptide margatoxin which inhibits the voltage-dependent potassium channels. It increases the time taken by a cell in order to execute an action potential in response to stimulation. It also affects nicotinic Ach-receptor in order to release norepinephrine which shows impacts on sympathetic control of cardiovascular function [67].
Potential against diabetes
Studies have revealed the anti-diabetic effects of scorpion toxins which also activate and generate ß-islets. Scorpion venom along with Chinese drugs is used to cure diabetes [68].
Potential against epilepsy
For the treatment of epilepsy, various antiepileptic drugs (AEDs) are used, which cause severe side effects, including teratogenesis, sedation, chronic toxicity, and cognitive impairment [69]. The specific peptides present in the venom of Chinese scorpion-Buthus martensi Karsch find applications as effective AEDs. The scorpion's entire body, particularly its tail, has been used in Chinese medicine to treat nervous disorders such as epilepsy, paralysis, and apoplexy [70]. The neurotoxins present in venom have a 3D backbone that helps them to bind efficiently for a long time. The venom of Leiurus quinquestriatus with alkaloid neurotoxins causes synergistic effects to regulate the action potential. When scorpion toxins are attached to receptors on dopaminergic neurons, then release of dopamine is observed, which may be effective for curing of Parkinson’s disease [71].
Potential against malaria
Peptides (present in scorpion venom) display anti-malarial properties (through their K+channels) and cell breakage by restricting activity at the carbon and nitrogen terminals. The first anti-malarial peptide was isolated from the venom of Pandinus imperator (family: Scorpionidae). This peptide can induce 98% of deaths in Plasmodium berghei at its sexual state; it causes 100% of death of Plasmodium falciparum parasites [72]. Mesobuthus eupeus contains antimalarial peptides namely meucin-13 and meucin-18 which induce cytolytic activity in various microbes, while meucin-24 and meucin-25 inhibit the activity of malarial parasites without causing any loss/impairment of normal mammalian cells [44].
CONCLUSION
A large number of neurotoxins are present in scorpion venom. These toxins have the ability to interrupt the normal functioning of multiple ion channels (Ca2+, Cl−, K+, and Na+) in excitable and non-excitable tissues. The scorpion venom finds an immense significance in the therapy of numerous diseases, including cancer, cardiovascular diseases, diabetes, AIDS, apoplexy, paralysis, epilepsy, malaria, measles, influenza H5N1, severe acute respiratory syndrome, Herpes simplex virus type 1 infection and diabetes etc with minimum or no side effects. It can induce anti-proliferation, apoptosis, cytotoxicity, and immunosuppressive effects that mainly inhibit the growth of disease-causing cells. The cell cycle, cell proliferation, and cell growth can all be influenced by various components of scorpion venom. Mucroporin-M is a component of venom which can be extracted from scorpion Lychas mucronatus; it is a good antiseptic agent for hand washes and mouthwashes due to its dual antimicrobial activity (antiviral and antibacterial). The computational studies may have a crucial role while developing ion channel drugs from venom peptides.
FUNDING
It is a review article and there were not funding sources.
AUTHORS CONTRIBUTIONS
All the authors have contributed equally.
CONFLICT OF INTERESTS
There is no conflict of interest between the authors for the manuscript.
REFERENCES
-
Ahn MY, Ryu KS, Lee YW, Kim YS. Cytotoxicity and L-amino acid oxidase activity of crude insect drugs. Arch Pharm Res. 2000;23(5):477-81. doi: 10.1007/BF02976576, PMID 11059827.
-
Petricevich VL. Effect of tityus serrulatus venom on cytokine production and the activity of murine macrophages. Mediators Inflamm. 2002;11(1):23-31. doi: 10.1080/09629350210308, PMID 11926592.
-
KV R. Study the assessment of poisoning cases in a rural tertiary care teaching hospital by a clinical pharmacist. Asian J Pharm Clin Res. 2012;5(2):138-41.
-
Ibrahim F, Sahlan M, Ginting MJ, Pratami DK, Hermansyah H, Wijanarko A. Isolation of protein from the spine venom of Pterois volitans found in the Indonesian ocean, using a heating process, for anticancer, antiretroviral, antibacterial, and antioxidant assays. Int J App Pharm. 2021;13Special Issue 2:53-8. doi: 10.22159/ijap.2021.v13s2.10.
-
Rajan SS, Subramanian P, Merlin JPJ. Snake venom-derived peptides as prospective pharmacological tools: recent trends. Int J Curr Pharm Sci. 2022;14(1):1-14. doi: 10.22159/ijcpr.2022v14i1.44106.
-
Bharadwa PV, Gogari PK. Possible action mechanism of bio-molecules extracted from snake venom as antivirals to inactivate novel corona virus. Crit Rev. 2020;10:589-91.
-
Sarkar S, Chakraverty R, Datta S, Ghosh A, Sarkar S. In vitro assays for neutralization of snake venom using herbal drugs: A creative. J Pharm Res. 2015;1(4):156-69.
-
Das Gupta SD, Debnath A, Saha A, Giri B, Tripathi G, Vedasiromoni JR. Indian black scorpion (Heterometrus bengalensis Koch) venom induced antiproliferative and apoptogenic activity against human leukemic cell lines U937 and K562. Leuk Res. 2007;31(6):817-25. doi: 10.1016/j.leukres.2006.06.004, PMID 16876244.
-
Chippaux JP, Goyffon M. Epidemiology of scorpionism: a global appraisal. Acta Trop. 2008;107(2):71-9. doi: 10.1016/j.actatropica.2008.05.021, PMID 18579104.
-
Al Asmari AK, Al Zahrani AG, Al Jowhary S, Arshaduddin M. Clinical aspects and frequency of scorpion stings in the Riyadh region of Saudi Arabia. Saudi Med J. 2012;33(8):852-8. PMID 22886117.
-
Al Asmari AK, Khan HA, Manthiri RA, Al Yahya KM, Al Otaibi KE. Effects of Echis pyramidum snake venom on hepatic and renal antioxidant enzymes and lipid peroxidation in rats. J Biochem Mol Toxicol. 2014;28(9):407-12. doi: 10.1002/jbt.21578, PMID 24888330.
-
Ozkan O, Filazi A. The determination of acute lethal dose-50 (LD50) levels of venom in mice, was obtained by different methods from scorpions, Androctonus crassicauda (Oliver 1807). Acta Parasitol Turc. 2004;28(1):50-3.
-
Ahmadi S, Knerr JM, Argemi L, Bordon KCF, Pucca MB, Cerni FA. Scorpion venom: detriments and benefits. Biomedicines. 2020;8(5):118. doi: 10.3390/biomedicines8050118, PMID 32408604.
-
Al-Asmari AK, Riyasdeen A, Islam M. Scorpion venom causes upregulation of p53 and downregulation of Bcl-xL and BID protein expression by modulating signaling proteins ERK1/2 and STAT3, and DNA damage in breast and colorectal cancer cell lines. Integr Cancer Ther. 2018;17(2):271-81. doi: 10.1177/1534735417704949, PMID 28438053.
-
El-Ghlban S, Kasai T, Shigehiro T, Yin HX, Sekhar S, Ida M. Chlorotoxin-Fc fusion inhibits the release of MMP-2 from pancreatic cancer cells. BioMed Res Int. 2014;2014:152659. doi: 10.1155/2014/152659, PMID 24511528.
-
Andreotti N, Jouirou B, Mouth L, Sabatier J. Comprehensive natural products II. Amsterdam: Elsevier; 2010.
-
Diaz Garcia A, Morier Diaz L, Frion Herrera Y, Rodriguez Sanchez H, Caballero-Lorenzo Y, Mendoza-Llanes D. In vitro anticancer effect of venom from Cuban scorpion Rhopalurus junceus against a panel of human cancer cell lines. J Venom Res. 2013;4:5-12. PMID 23946884.
-
Gomes A, Bhattacharjee P, Mishra R, Biswas AK, Dasgupta SC, Giri B. Anticancer potential of animal venoms and toxins. Indian J Exp Biol. 2010;48(2):93-103. PMID 20455317.
-
Petricevich VL, Navarro LB, Possani LD. Therapeutic use of scorpion venom. Mol ASp Inflamm. 2013;9:209-31.
-
Oukkache N, Chgoury F, Lalaoui M, Cano AA, Ghalim N. Comparison between two methods of scorpion venom milking in Morocco. J Venom Anim Toxins Incl Trop Dis. 2013;19(1):5. doi: 10.1186/1678-9199-19-5, PMID 23849043.
-
Ortiz E, Gurrola GB, Schwartz EF, Possani LD. Scorpion venom components as potential candidates for drug development. Toxicon. 2015;93:125-35. doi: 10.1016/j.toxicon.2014.11.233, PMID 25432067.
-
Santibanez Lopez CE, Possani LD. Overview of the Knottin scorpion toxin-like peptides in scorpion venoms: insights on their classification and evolution. Toxicon. 2015;107(B):317-26. doi: 10.1016/j.toxicon.2015.06.029, PMID 26187850.
-
Ma Y, Zhao R, He Y, Li S, Liu J, Wu Y. Transcriptome analysis of the venom gland of the scorpion Scorpiops jendeki: implication for the evolution of the scorpion venom arsenal. BMC Genomics. 2009;10(1):290. doi: 10.1186/1471-2164-10-290, PMID 19570192.
-
Almaaytah A, Albalas Q. Scorpion venom peptides with no disulfide bridges: a review. Peptides. 2014;51:35-45. doi: 10.1016/j.peptides.2013.10.021, PMID 24184590.
-
Possani LD, Merino E, Corona M, Bolivar F, Becerril B. Peptides and genes coding for scorpion toxins that affect ion channels. Biochimie. 2000;82(9-10):861-8. doi: 10.1016/s0300-9084(00)01167-6, PMID 11086216.
-
Cestele S, Catterall WA. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie. 2000;82(9-10):883-92. doi: 10.1016/s0300-9084(00)01174-3, PMID 11086218.
-
Quintero Hernandez V, Jimenez Vargas JM, Gurrola GB, Valdivia HH, Possani LD. Scorpion venom components that affect ion-channels function. Toxicon. 2013;76:328-42. doi: 10.1016/j.toxicon.2013.07.012, PMID 23891887.
-
Fry BG. From genome to ”venome”: molecular origin and evolution of the snake venom proteome inferred from phylogenetic analysis of toxin sequences and related body proteins. Genome Res. 2005;15(3):403-20. doi: 10.1101/gr.3228405, PMID 15741511.
-
Chen R, Chung SH. Computational studies of venom peptides targeting potassium channels. Toxins. 2015;7(12):5194-211. doi: 10.3390/toxins7124877, PMID 26633507.
-
Bagal SK, Brown AD, Cox PJ, Omoto K, Owen RM, Pryde DC. Ion channels as therapeutic targets: a drug discovery perspective. J Med Chem. 2013;56(3):593-624. doi: 10.1021/jm3011433, PMID 23121096.
-
Chandy KG, Wulff H, Beeton C, Pennington M, Gutman GA, Cahalan MD. K+channels as targets for specific immunomodulation. Trends Pharmacol Sci. 2004;25(5):280-9. doi: 10.1016/j.tips.2004.03.010, PMID 15120495.
-
Knapp O, McArthur JR, Adams DJ. Conotoxins targeting neuronal voltage-gated sodium channel subtypes: potential analgesics? Toxins. 2012;4(11):1236-60. doi: 10.3390/toxins4111236, PMID 23202314.
-
Shi J, He HQ, Zhao R, Duan YH, Chen J, Chen Y. Inhibition of martentoxin on neuronal BK channel subtype (alpha+beta4): implications for a novel interaction model. Biophys J. 2008;94(9):3706-13. doi: 10.1529/biophysj.107.122150, PMID 18199674.
-
Shieh CC, Coghlan M, Sullivan JP, Gopalakrishnan M. Potassium channels: molecular defects, diseases, and therapeutic opportunities. Pharmacol Rev. 2000;52(4):557-94. PMID 11121510.
-
Tenenholz TC, Klenk KC, Matteson DR, Blaustein MP, Weber DJ. Structural determinants of scorpion toxin affinity: the charybdotoxin (alpha-KTX) family of K(+)-channel blocking peptides. Rev Physiol Biochem Pharmacol. 2000;140:135-85. doi: 10.1007/BFb0035552, PMID 10857399.
-
Holaday Jr SK, Martin BM, Fletcher Jr PL, Krishna NR. NMR solution structure of butantoxin. Arch Biochem Biophys. 2000;379(1):18-27. doi: 10.1006/abbi.2000.1858, PMID 10864437.
-
Cao Z, Di Z, Wu Y, Li W. Overview of scorpion species from China and their toxins. Toxins. 2014;6(3):796-815. doi: 10.3390/toxins6030796, PMID 24577583.
-
Yu FH, Catterall WA. Overview of the voltage-gated sodium channel family. Genome Biol. 2003;4(3):207. doi: 10.1186/gb-2003-4-3-207, PMID 12620097.
-
Mander L, Liu HW. Comprehensive natural products II: Chemistry and biology. Elsevier; 2010.
-
Matsushita N, Miyashita M, Sakai A, Nakagawa Y, Miyagawa H. Purification and characterization of a novel short-chain insecticidal toxin with two disulfide bridges from the venom of the scorpion Liocheles australasiae. Toxicon. 2007;50(6):861-7. doi: 10.1016/j.toxicon.2007.06.014, PMID 17681581.
-
Ding J, Chua PJ, Bay BH, Gopalakrishnakone P. Scorpion venoms as a potential source of novel cancer therapeutic compounds. Exp Biol Med (Maywood). 2014;239(4):387-93. doi: 10.1177/1535370213513991, PMID 24599885.
-
Becchetti A. Ion channels and transporters in cancer. 1. Ion channels and cell proliferation in cancer. Am J Physiol Cell Physiol. 2011;301(2):C255-65. doi: 10.1152/ajpcell.00047.2011, PMID 21430288.
-
Ahluwalia S, Shah N. Animal venom for treating breast cancer. Int J Pharm Pharm Sci. 2014;6(9):24-30.
-
Ghosh A, Roy R, Nandi M, Mukhopadhyay A. Scorpion venom–toxins that aid in drug development: a review. Int J Pept Res Ther. 2019;25(1):27-37. doi: 10.1007/s10989-018-9721-x, PMID 32214927.
-
Lorenzo LD. Cancer pain management with a venom of blue scorpion endemic in cuba, called rhopalurus junceus ”Escozul”. Open Cancer J. 2012;5(1):1-2. doi: 10.2174/1874079001205010001.
-
D’Suze G, Rosales A, Salazar V, Sevcik C. Apoptogenic peptides from tityus discrepans scorpion venom acting against the SKBR3 breast cancer cell line. Toxicon. 2010;56(8):1497-505. doi: 10.1016/j.toxicon.2010.09.008, PMID 20888852.
-
Mishal R, Tahir HM, Zafar K, Arshad M. Anticancerous applications of scorpion venom. Int J Biol Pharm Res. 2013;4:356-60.
-
Zargan J, Umar S, Sajad M, Naime M, Ali S, Khan HA. Scorpion venom (Odontobuthus doriae) induces apoptosis by depolarization of mitochondria and reduces S-phase population in human breast cancer cells (MCF-7). Toxicol In Vitro. 2011;25(8):1748-56. doi: 10.1016/j.tiv.2011.09.002, PMID 21945044.
-
Chaisakul J, Hodgson WC, Kuruppu S, Prasongsook N. Effects of animal venoms and toxins on hallmarks of cancer. J Cancer. 2016;7(11):1571-8. doi: 10.7150/jca.15309, PMID 27471574.
-
Lyons SA, O’Neal J, Sontheimer H. Chlorotoxin, a scorpion-derived peptide, specifically binds to gliomas and tumors of neuroectodermal origin. Glia. 2002;39(2):162-73. doi: 10.1002/glia.10083, PMID 12112367.
-
Veiseh M, Gabikian P, Bahrami SB, Veiseh O, Zhang M, Hackman RC. Tumor paint: a chlorotoxin: Cy5. 5 bioconjugates for intraoperative visualization of cancer foci. Cancer Res. 2007;67(14):6882-8. doi: 10.1158/0008-5472.CAN-06-3948, PMID 17638899.
-
Feng L, Gao R, Gopalakrishnakone P. Isolation and characterization of a hyaluronidase from the venom of Chinese red scorpion Buthus martensi. Comp Biochem Physiol C Toxicol Pharmacol. 2008;148(3):250-7. doi: 10.1016/j.cbpc.2008.06.003, PMID 18611448.
-
Sariego J. Breast cancer in the young patient. Am Surg. 2010;76(12):1397-400. doi: 10.1177/000313481007601226, PMID 21265355.
-
Feng Y, Broder CC, Kennedy PE, Berger EA. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science. 1996;272(5263):872-7. doi: 10.1126/science.272.5263.872, PMID 8629022.
-
Deng H, Liu R, Ellmeier W, Choe S, Unutmaz D, Burkhart M. Identification of a major co-receptor for primary isolates of HIV-1. Nature. 1996;381(6584):661-6. doi: 10.1038/381661a0, PMID 8649511.
-
Chen Y, Cao L, Zhong M, Zhang Y, Han C, Li Q. Anti-HIV-1 activity of a new scorpion venom peptide derivative Kn2-7. PLOS ONE. 2012;7(4):e34947. doi: 10.1371/journal.pone.0034947, PMID 22536342.
-
Gopinath SC, Hayashi K, Kumar PK. Aptamer binds to the gD protein of herpes simplex virus 1 and efficiently inhibits viral entry. J Virol. 2012;86(12):6732-44. doi: 10.1128/JVI.00377-12, PMID 22514343.
-
Hill JM, Gebhardt BM, Wen R, Bouterie AM, Thompson HW, O’Callaghan RJ. Quantitation of herpes simplex virus type 1 DNA and latency-associated transcripts in rabbit trigeminal ganglia demonstrates a stable reservoir of viral nucleic acids during latency. J Virol. 1996;70(5):3137-41. doi: 10.1128/JVI.70.5.3137-3141.1996, PMID 8627793.
-
Hong W, Li T, Song Y, Zhang R, Zeng Z, Han S. Inhibitory activity and mechanism of two scorpion venom peptides against herpes simplex virus type 1. Antiviral Res. 2014;102:1-10. doi: 10.1016/j.antiviral.2013.11.013, PMID 24315793.
-
van Soest H, Renooij W, van Erpecum KJ. Clinical and basal aspects of anemia during antiviral therapy for hepatitis C. Ann Hepatol. 2009;8(4):316-24. doi: 10.1016/S1665-2681(19)31744-2, PMID 20009130.
-
Sleeman K, Stein DA, Tamin A, Reddish M, Iversen PL, Rota PA. Inhibition of measles virus infections in cell cultures by peptide-conjugated morpholino oligomers. Virus Res. 2009;140(1-2):49-56. doi: 10.1016/j.virusres.2008.10.018, PMID 19059443.
-
Dai C, Ma Y, Zhao Z, Zhao R, Wang Q, Wu Y. Mucroporin, the first cationic host defense peptide from the venom of Lychas mucronatus. Antimicrob Agents Chemother. 2008;52(11):3967-72. doi: 10.1128/AAC.00542-08, PMID 18779362.
-
Yan R, Zhao Z, He Y, Wu L, Cai D, Hong W. A new natural α-helical peptide from the venom of the heteromerous scorpion petersii kills HCV. Peptides. 2011;32(1):11-9. doi: 10.1016/j.peptides.2010.10.008, PMID 20950663.
-
Li Q, Zhao Z, Zhou D, Chen Y, Hong W, Cao L. Virucidal activity of a scorpion venom peptide variant mucroporin-M1 against measles, SARS-CoV and influenza H5N1 viruses. Peptides. 2011;32(7):1518-25. doi: 10.1016/j.peptides.2011.05.015, PMID 21620914.
-
Lu X, Lu D, Scully MF, Kakkar VV. Integrins in drug targeting-RGD templates in toxins. Curr Pharm Des. 2006;12(22):2749-69. doi: 10.2174/138161206777947713, PMID 16918409.
-
McLane MA, Joerger T, Mahmoud A. Disintegrins in health and disease. Front Biosci. 2008;13(1):6617-37. doi: 10.2741/3177, PMID 18508683.
-
Cheong A, Li J, Sukumar P, Kumar B, Zeng F, Riches K. Potent suppression of vascular smooth muscle cell migration and human neointimal hyperplasia by KV1. 3 Channel blockers. Cardiovasc Res. 2011;89(2):282-9. doi: 10.1093/cvr/cvq305, PMID 20884640.
-
Xie J, Herbert TP. The role of mammalian target of rapamycin (mTOR) in the regulation of pancreatic β-cell mass: implications in the development of type-2 diabetes. Cell Mol Life Sci. 2012;69(8):1289-304. doi: 10.1007/s00018-011-0874-4, PMID 22068611.
-
Raza M, Shaheen F, Choudhary MI, Sombati S, Rafiq A, Suria A. Anticonvulsant activities of ethanolic extract and aqueous fraction isolated from Delphinium denudatum. J Ethnopharmacol. 2001;78(1):73-8. doi: 10.1016/s0378-8741(01)00327-0, PMID 11585691.
-
Villetti G, Bregola G, Bassani F, Bergamaschi M, Rondelli I, Pietra C. Preclinical evaluation of CHF3381 as a novel antiepileptic agent. Neuropharmacology. 2001;40(7):866-78. doi: 10.1016/s0028-3908(01)00026-0, PMID 11378157.
-
Tobassum S, Tahir HM, Arshad M, Zahid MT, Ali S, Ahsan MM. Nature and applications of scorpion venom: an overview. Toxin Rev. 2020;39(3):214-25. doi: 10.1080/15569543.2018.1530681.
-
Ramirez KL, Jimenez Vargas JM. Scorpine-like peptides. Single Cell Biol. 2016;5(2). doi: 10.4172/2168-9431.1000138.
-
Xu J, Zhang X, Guo Z, Yan J, Yu L, Li X. Orthogonal separation and identification of long-chain peptides from the scorpion Buthus martensi Karsch venom by using two-dimensional mixed-mode reversed phase-reversed phase chromatography coupled to tandem mass spectrometry. Analyst. 2013;138(6):1835-43. doi: 10.1039/c2an36704a, PMID 23373063.
-
Possani LD, Becerril B, Delepierre M, Tytgat J. Scorpion toxins specific for Na+‐channels. Eur J Biochem. 1999;264(2):287-300. doi: 10.1046/j.1432-1327.1999.00625.x, PMID 10491073.
-
CN104193813A C. Sep purif method scorpion venom polypeptide use thereof; 2014.
-
Cordeiro FA, Amorim FG, Anjolette FA, Arantes EC. Arachnids of medical importance in Brazil: main active compounds present in scorpion and spider venoms and tick saliva. J Venom Anim Toxins Incl Trop Dis. 2015;21:24. doi: 10.1186/s40409-015-0028-5, PMID 26273285.
-
Hmed B, Serria HT, Mounir ZK. Scorpion peptides: potential use for new drug development. J Toxicol. 2013;2013:958797. doi: 10.1155/2013/958797, PMID 23843786.
-
Pennington MW, Czerwinski A, Norton RS. Peptide therapeutics from venom: current status and potential. Bioorg Med Chem. 2018;26(10):2738-58. doi: 10.1016/j.bmc.2017.09.029, PMID 28988749.
-
Oukkache N, Chgoury F, Lalaoui M, Cano AA, Ghalim N. Comparison between two methods of scorpion venom milking in Morocco. J Venom Anim Toxins Incl Trop Dis. 2013;19(1):5. doi: 10.1186/1678-9199-19-5, PMID 23849043.
-
Norton RS, Chandy KG. Venom-derived peptide inhibitors of voltage-gated potassium channels. Neuropharmacology. 2017;127:124-38. doi: 10.1016/j.neuropharm.2017.07.002, PMID 28689025.
-
Garcia Calvo M, Leonard RJ, Novick J, Stevens SP, Schmalhofer W, Kaczorowski GJ. Purification, characterization, and biosynthesis of margatoxin, a component of Centruroides margaritatus venom that selectively inhibits voltage-dependent potassium channels. Journal of Biological Chemistry. 1993;268(25):18866-74. doi: 10.1016/S0021-9258(17)46707-X, PMID 8360176.
-
Koo GC, Blake JT, Talento A, Nguyen M, Lin S, Sirotina A. Blockade of the voltage-gated potassium channel Kv1. 3 inhibits immune responses in vivo. The Journal of Immunology. 1997;158(11):5120-8. PMID 9164927.
-
Lebrun B, Romi-Lebrun R, Martin Eauclaire MF, Yasuda A, Ishiguro M, Oyama Y. A four-disulfide-bridged toxin, with high affinity towards voltage-gated K+channels, isolated from Heterometrus spinnifer (Scorpionidae) venom. Biochemical J. 1997;328(1):321-7. doi: 10.1042/bj3280321, PMID 9359871.
-
Regaya I, Beeton C, Ferrat G, Andreotti N, Darbon H, De Waard M. Evidence for domain-specific recognition of SK and Kv channels by MTX and HsTx1 scorpion toxins. Journal of Biological Chemistry. 2004;279(53):55690-6. doi: 10.1074/jbc.M410055200, PMID 15498765.
-
Chandy KG, Norton RS. Peptide blockers of Kv1. 3 Channels in T cells as therapeutics for autoimmune disease. Current Opinion in Chem Biology. 2017;38:97-107. doi: 10.1016/j.cbpa.2017.02.015, PMID 28412597.
-
Yoshimoto Y, Miyashita M, Abdel Wahab M, Sarhan M, Nakagawa Y, Miyagawa H. Isolation and characterization of insecticidal toxins from the venom of the North African Scorpion, Buthacus leptochelys. Toxins. 2019;11(4):236. doi: 10.3390/toxins11040236, PMID 31027216.
-
Erdeş E, Dogan TS, Coşar I, Danışman T, Kunt KB, Seker T. Characterization of Leiurus abdullahbayrami (Scorpiones: Buthidae) venom: peptide profile, cytotoxicity and antimicrobial activity. Journal of Venom Anim Toxins Incl Trop Dis. 2014;20(1):1-8:48. doi: 10.1186/1678-9199-20-48, PMID 25414725.
-
Smith JJ, Hill JM, Little MJ, Nicholson GM, King GF, Alewood PF. Unique scorpion toxin with a putative ancestral fold provides insight into the evolution of the inhibitor cystine knot motif. Proceedings of the National Academy of Sciences USA. 2011;108(26):10478-83. doi: 10.1073/pnas.1103501108, PMID 21670253.
-
Ji YH, Wang WX, Ye JG, He LL, Li YJ, Yan YP. Martentoxin, a novel K+‐channel‐blocking peptide: purification, cDNA and genomic cloning, and electrophysiological and pharmacological characterization. Journal of Neurochemistry. 2003;84(2):325-35. doi: 10.1046/j.1471-4159.2003.01516.x, PMID 12558995.